https://barryhisblog.blogspot.com/p/on-failures-of-mathematical-anti.html
I'm a formally trained and experienced teacher of mathematics. I trained at a Christian-founded college of education where they have no problems with evolutionary science. https://barryhisblog.blogspot.com/p/arent-you-just-plagiarist-why-dont-you.html
The way in which probability, a branch of mathematics, is, possibly deliberately and knowingly abused by creationists and intelligent designer spotters really bugs me. Not only do they abuse the mathematics, but also gullible followers, who, as everyone knows, include lay people in general who struggle with probability, often finding it counter-intuitive, despite the fact that it is sound.
I produced a routine that illustrates a very basic error in probability theory, an error which many creationists make and cling to because they think it supports their position. It does not. https://barryhisblog.blogspot.com/p/probability-emo.html
I was therefore very grateful to have this book recommended to me. I shall be developing this page further, delving into Jason's book.
And here is Jerry Coyne's article on Jason's book. https://whyevolutionistrue.com/2022/06/02/the-intellectual-vacuity-of-mathematical-arguments-against-evolution/

https://www.amazon.com/Failures-Mathematical-Anti-Evolutionism-Jason-Rosenhouse-ebook/dp/B0B523NRCB
Jason Rosenhouse is a professor of mathematics at James Madison University.
Anti-scientific misinformation has become a serious problem on many fronts, including vaccinations and climate change. One of these fronts is the persistence of anti-evolutionism, which has recently been given a superficially professional gloss in the form of the intelligent design movement. Far from solely being of interest to researchers in biology, anti-evolutionism must be recognized as part of a broader campaign with a conservative religious and political agenda. Much of the rhetorical effectiveness of anti-evolutionism comes from its reliance on seemingly precise mathematical arguments. This book, the first of its kind to be written by a mathematician, discusses and refutes these arguments. Along the way, it also clarifies common misconceptions about both biology and mathematics. Both lay audiences and professionals will find the book to be accessible and informative.
"If you want to convince an audience of a falsehood, an excellent strategy is to bamboozle them with mathematical jargon that they don't understand. If your incomprehensible equations seem to prove what the audience wants to believe anyway, you'll earn a standing ovation. This has been a favorite tactic of anti-evolutionists. Jason Rosenhouse is a good mathematician but, unlike many mathematicians, he is superbly gifted in the art of translating mathematical arguments into words. He mercilessly exposes creationist abuses of mathematics, using language that non-mathematicians can pleasurably follow. As a bonus along the way, he is an excellent mathematics teacher, patiently explaining each point before revealing the abuse of it. I especially appreciated his deft use of analogies. He is thoroughly well read in the bogus literature of creationism and 'intelligent design,' as well as in the biological literature. We have here a superb book, lucid, knowledgeable, wise, and very necessary."
Richard Dawkins FRS, Emeritus Professor of the Public Understanding of Science, University of Oxford
"A little mathematics can be a dangerous thing when it is used as a rhetorical weapon in a political and religious battle. In this incisive and crystal clear book, Jason Rosenhouse shows that the arguments of mathy creationists are like the proverbial spherical cow and unflightworthy bumblebee: sophistical, wrongheaded, and out of touch with biological reality." Steven Pinker, Johnstone Professor of Psychology, Harvard University, and the author of How the Mind Works and Rationality
"Creationists beware! Jason Rosenhouse is here to debunk your perversions of probability theory, your tortured thermodynamics, and your insults to information theory. And the rest of us will learn a lot about evolution, mathematics, and their interrelations from the expert guidance offered in The Failures of Mathematical Anti-Evolutionism." Glenn Branch, Deputy Director, National Center for Science Education
"Written by a mathematician who has paid close attention to mathematical arguments against evolution, this is a beautifully written and careful refutation of those arguments. Jason Rosenhouse covers. arguments based on improbability, information, and thermodynamics, and he does so with clear explanations that will be accessible to anyone who takes science seriously. A wonderful achievement." Joe Felsenstein, University of Washington
So here are my comments and observations regarding the core thrust of Rosenhouse's book, chapter 5, part 5.5, beginning on page 124.
https://barryhisblog.blogspot.com/p/here-be-dragons-searching-large-phase.html
5.5 THE BASIC ARGUMENT FROM IMPROBABILITY
We have noted that they start with a track one argument that evolution is asking us to believe that something very improbable has occurred. Their various track two arguments are then attempts to state precisely what that something is. For this purpose, they often take aim at the specificity of proteins, and we will consider some of their efforts in this regard in the present section.
We start with an especially clear example, written by David Foster, an ID proponent. His argument involves hemoglobin, a pro- tein that transports oxygen through the blood of humans and other vertebrates.
Hemoglobin is a complex molecule possessing an elaborate, three-dimensional structure. However, Foster's argument treats it as a simple string of 574 amino acids. This is a crude oversimplification, but we can accept it for the sake of argument. Our goal is to expose the fallacious mathematical reasoning underlying the argument, as opposed to obsessing about every biological detail.
There are twenty kinds of amino acids in total. Foster begins his argument by listing the frequency with which each amino acid appears in hemoglobin. For example, the amino acid glycine appears 36 times, alanine appears 68 times, and so on. He now writes:
The specificity of hemoglobin is described by the improbability of the specific amino acid sequence occurring by random chance. Such specificity is capable of exact calculation in the permutation formula:
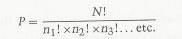
where N is the total number of amino acids in hemoglobin (574); n1, etc. are the number of separate kinds of amino acids; and ! means that the given separate numbers are subjected to "factorial" expansion. Thus: 5!= 5 x 4 × 3 × 2.
In the case of hemoglobin, ... the specific numerical value of the solution is P = 10654. This is an immense number, 10 multiplied by itself 654 times. ... Thus we can state that the improbability of hemoglobin occurring by random selection can be represented by the infinitely small number 10-654, which means 10 divided by itself 654 times: as near to zero as one could consider. ... This raises the question as to whether such very low probabilities are of a miraculous nature when they occur in factual situations such as the protein hemoglobin - whether such extremely improbable events are relevant to the question "Does God exist?"
For the sake of accuracy, we should note that 10-654 does not actually mean to divide 10 by itself 654 times. Rather, it means to divide 1 by 10 a total of 654 times. This is equivalent to dividing 10 by itself 653 times.
Arguments of this general sort are ubiquitous in anti- evolutionist literature. Foster's article was published in a mainstream American periodical called The Saturday Evening Post, and I am sure its generally conservative readership was delighted by the superficial precision of his argument. I used Roth's example because he presented an easily quotable version of the argument, but most young-Earth creationist books present some version of it.
Regardless of the precise variation, the underlying logic of the argument is always the same:
1. Identify a complex biological structure, such as a specific gene or protein. 2. Model its evolution as a process of randomly selecting one item from a very large space of equiprobable possibilities.
3. Use elementary combinatorics to determine the size of the space, which we shall call S.
4. Conclude that the probability of the structure having evolved by chance
is 1/S, and assert that this is too small for evolution to be plausible.
I shall refer to this as the Basic Argument from Improbability (BAI). What can be said in reply?
Let us recast the BAI in the language of Section 5.2. We will assume the argument is being applied to a protein. In this case, our probability space consists of the set of proteins of a certain length. The BAI equips this space with the uniform distribution, meaning, recall, that we assume that any individual protein is as likely to occur as any other. Evolving a specific protein is then modeled as selecting one point at random from this space. Since the space is vast, the number of attempts needed to achieve success is prohibitive, even
5.5 THE BASIC ARGUMENT FROM IMPROBABILITY 127
given geological time scales with which to work. At this point you assert that evolution has been disproved and call it a day.
However, you will have noticed that a critical part of the evolutionary process has been left out of this model. Biologists do not claim that the proteins found in modern organisms arose in one step by random selection from the space of all theoretical possibilities. The actual claim is that they arose in a gradual, step-by-step manner, as random variations were passed through the sieve of natural selection. Moreover, as we have noted, the problem is never to explain how some modern protein arose from scratch, but is rather to explain how the modern protein evolved from some more ancient protein, one that was possibly simpler and more rudimentary than the modern form.
Recall also our discussion from Section 4.2. Evolution finds its
ultimate starting point at the origin of life, which we can think of as situating life at some specific point in protein space. As the process plays out, we do not search protein space as a whole, but instead carry out a sequence of local searches in the neighborhood of the point at which we happen to find ourselves. We noted that the vastness of the space as a whole is irrelevant because we only search a tiny fraction of it. This is illustrated in Figure 5.2.
We can now point to the first serious error with the BAI: it equipped its probability space with the uniform distribution, which is terribly unrealistic. Parents are very unlikely to have offspring with a protein far away from theirs. It is much more likely that the offspring has a protein that is local to the parents. Therefore, proteins near to our starting point are far more likely to be sampled than are proteins that are far away. The correct distribution would assign a probability close to 0 for most of the space since most proteins will be very far away from our starting point - and then a very high probability to proteins that are nearby.
This leads us to the second serious problem with the BAI: it ignores the role of natural selection in affecting the probability of finding certain proteins. Suppose that some nearby proteins represent improvements over our current state, while other nearby proteins are
128
5 PROBABILITY THEORY
Here be dragons.
Here be dragons.
Origin of life
FIGURE 5.2 Searching protein space. The solid black dot at the intersection of the axes represents the origin of life. The other solid black dots represent protein stepping stones connecting an ancient protein to more modern forms. The gray circles show that evolution only examines the local area near an already existing protein and completely disregards most of the space.
harmful. Natural selection will then ensure that the improvements are more likely to be represented in future generations than are the harmful proteins. Mathematically speaking, we could say that the effect of selection is to dramatically shift the probability distribution toward favorable proteins and away from harmful ones. If there is a path of steady improvement connecting the rudimentary protein to its modern descendant, then natural selection will preserve our progress along that path. It will prevent us from moving backward while we wait for the next improvement.
We can dramatize these issues with an analogy. Suppose you and a friend are in the downtown area of a major American city, and you both decide you want a slice of pizza. You pick a direction and
5.5 THE BASIC ARGUMENT FROM IMPROBABILITY 129
start walking. Within just two blocks you find a pizza parlor. Your friend now says, "Incredible! The surface of the Earth is enormous, and almost none of it is covered with pizza parlors. Yet somehow we were able to find one of the few places on Earth that has a pizza parlor. How can you explain something so remarkable?"
In this context, the error is obvious. The surface area of the earth is irrelevant because we only needed to search the tiny portion of it near our current location. And while pizza parlors are rare on the surface of the earth generally, they are extremely common in the downtown areas of major American cities. The BAI is guilty of precisely the same oversights, except applied to protein space rather
than to the surface of the earth.
A more recent version of this fallacy occurs in the book Dar- win's Doubt, written by ID proponent Stephen Meyer. Referring to the Wistar conference we discussed in Chapter 4, he writes:
And that was the problem, as the Wistar skeptics saw it: random mutation must do the work of composing new genetic information, yet the sheer number of possible nucleotide base or amino-acid combinations (i.e. the size of the combinatorial
"space") associated with a single gene or protein of even modest length rendered the probability of random assembly prohibitively small. For every sequence of amino acids that generates a functional protein, there are a myriad of other combinations that don't. As the length of the required protein grows, the number of possible amino-acid combinations mushrooms exponentially. As this happens, the probability of ever stumbling by random
mutation onto a functional sequence rapidly diminishes.
(Meyer 2013, 173)
The logic underlying this argument fits our bullet-point list perfectly, and Meyer's argument fails for the reasons we have already enumerated. The probability space implied by his argument is entirely unrealistic biologically since it includes no role for natural
selection.
130 5 PROBABILITY THEORY
However, unlike Foster and Roth, Meyer is too sophisticated to ignore natural selection altogether. Just prior to our previous quote, he presents his reason for discounting it:
Clearly, natural selection plays a crucial role in this process. Favorable mutations are passed on; unfavorable mutations are weeded out. Nevertheless, the process can only select variations in the genetic text that mutations have first produced. For this
reason, evolutionary biologists typically recognize that mutation, not natural selection, provides the source of variation and innovation in the evolutionary process. As evolutionary biologists Jack King and Thomas Jukes put it in 1969, "Natural selection is the editor, rather than the composer, of the genetic message."
(Meyer 2013, 173)
This is a remarkable paragraph. One would have thought it obvious that mutation and selection both play a crucial role in the process of evolutionary innovation. According to evolutionary theory, complex, functional structures arise through the interplay of mutation and selection. It does not make sense to say that one or the other is the ultimate source of the adaptation. It is fine to say that mutation provides the ultimate source of variation, but it is flatly wrong to say it is the source of innovation. Continuing with the analogy of writers and editors, mutation is like a sloppy, self-indulgent writer who buries a few pages of insightful writing within hundreds of pages of unpublishable nonsense. The editor then plays a crucial role in the process by separating the few worthy pages from the mountains of chaff.
Consider an analogy. Imagine a child learning to play chess. He plays many practice games with his coach, who dutifully critiques his moves and shows him better ways of playing. Years later the child becomes a master-strength player. Meyer's argument is tantamount to saying that the coach played no role in the process of the student becoming a master. After all, the coach had nothing to critique until the child actually made some moves at the board. In the analogy,
5.5 THE BASIC ARGUMENT FROM IMPROBABILITY 131
mutation is like the child making moves and natural selection is like the coach passing judgment on them. And just as the child's moves and the coach's commentary were both essential to turning the child into a master, so too are mutation and selection both essential to the process of biological innovation.
Summarizing, we have here another instance of evolution's crit- ics failing to take into consideration the probabilistic and geometric structure of protein space. Specifically, the probability distribution appropriate to protein space is highly nonuniform, meaning that some proteins are far more likely to be found in organisms than others. And our movement through protein space is highly constrained by its geometric structure since each generation almost exclusively investigates points that are near to the generation before. The BAI ignores both of these points and can be dismissed for these reasons.
There is also a third serious problem with the BAI: It treats low probability, all by itself, as a refutation of evolution and confirmation of design. The problem with this approach is that extremely improba- ble events happen all the time. We have previously mentioned the old saying that million to one odds happen eight times a day in New York City. The biological analog is this: The course of evolution is affected by so many chance events that any specific modern outcome of the process could be extremely low, but this does not make us suspect design, because something had to happen. Therefore, some additional argument is needed to go from low probability to a conclusion of design. We need to argue not simply that, starting from the origin of life, a given protein was unlikely to have evolved, but also that this is the sort of improbability that needs a special explanation. Proponents of the BAI do not supply that additional argument.
The BAI has the great virtue of computational tractability. By reducing probability to combinatorics, its proponents are able to carry out actual calculations to produce the desired small numbers. However, we have seen that the mathematical model on which the BAI relies is far too unrealistic to produce meaningful results. Evolution does not proceed by randomly selecting complex structures
1
132 5 PROBABILITY THEORY
from spaces of equiprobable possibilities, but instead builds them gradually by accumulating many small changes, a process we can refer to as "cumulative selection."
Including a role for cumulative selection in the model would make it all but impossible to carry out meaningful calculations. The number of variables influencing the probabilistic structure of protein space is enormous. It is effectively impossible to identify them all, much less to assign appropriate numbers to them.
Thus, anyone seeking to use probability theory to refute evolu- tion must deal seriously with these two questions:
How do we preserve the computational tractability of the BAI while working within a biologically reasonable model?
• How do we justify the conclusion that evolution has been refuted simply because a particular mathematical model tells us that an event of low probability has occurred?
The remainder of this chapter will discuss the main attempts from ID proponents to circumvent these issues. As we shall see, they are not successful in doing so.
writes:
When trying to explain something, we employ three broad modes of explanation: necessity, chance and design. As a criterion for detecting design, specified complexity enables us to decide which of these modes of explanation apply. It does that by answering three questions about the thing we are trying to explain: Is it
are you a theistic evolution person
ReplyDeleteAtheist, but not anti-theist. Anti-creationist.
Delete